Inception : la manipulation des souvenirs pendant le sommeil pour mieux comprendre la mémoire
Le sommeil et la mémoire sont deux des fonctions les plus fondamentales pour la survie dans le règne animal. Chacun de nous peut sentir l’importance du sommeil sur son quotidien lorsqu’il fait l’expérience de son manque. Ce déficit de sommeil entraine l’apparition d’un sentiment de fatigue et affecte les fonctions exécutives, les processus attentionnels et la mémoire. Il est encore très difficile de comprendre précisément, d’un point de vue scientifique, les bases cérébrales de cette fatigue et on ne connait guère plus les mécanismes qui expliquent les effets bénéfiques du sommeil sur l’attention. En revanche, depuis les années 90, on a pu identifier certains processus neuronaux liés à l’amélioration de la mémoire pendant le sommeil, ce qui a fortement dynamisé la recherche scientifique dans ce domaine. Ces découvertes ont été largement commentées dans les médias et il nous parait ici capital de reprendre leurs bases scientifiques et de les replacer dans leur contexte. Nous commencerons donc par une description sommaire du support biologique de la mémoire, ou plutôt des mémoires, avant de détailler les deux grandes hypothèses du rôle du sommeil dans la mémoire. Nous terminerons notre propos en exposant les différents moyens mis à notre disposition pour améliorer les bienfaits du sommeil, sa possible utilisation pour le traitement du stress post-traumatique et nous verrons comment comprendre les mécanismes du sommeil nous aide finalement à mieux comprendre ceux de l’apprentissage durant l’éveil.
Le patient HM et les différents types de mémoire
Après les observations du début du 20ème siècle faites par Korsakov, Calparede et Ebbinghaus, c’est sûrement l’étude d’un unique patient, Henry Gustav Molaison, plus connu sous le nom de patient H.M., qui a permis de lever plus clairement le voile sur les grands mystères de la mémoire. Henry Molaison souffrait d’une forme d’épilepsie résistante aux traitements pharmacologiques. Les chirurgiens ont donc utilisé la dernière option disponible à l’époque : l’ablation du foyer de l’épilepsie. Ce dernier était localisé à côté d’une petite structure du cerveau appelée « hippocampe ». Bien que guéri de ses crises d’épilepsies, l’opération ne fut pas un complet succès puisqu’elle causa à ce patient une profonde amnésie : il est devenu incapable de former de nouveaux souvenirs (amnésie antérograde), démontrant ainsi l’importance de l’hippocampe dans la mémorisation. Mais ce déficit purement mnésique montra également que la mémoire est séparée des autres fonctions cognitives. Henry Molaison était en effet capable de réaliser la plupart des activités tout à fait normalement mais ne pouvait pas s’en souvenir au-delà d’une période de quelques minutes.
De manière surprenante, l’ablation de l’hippocampe entraîna des déficits de mémoire pour les souvenirs factuels ou les évènements de sa vie, mais laissa intactes d’autres formes de mémoire comme les conditionnements simples ou les apprentissages moteur. Cette dichotomie aboutit finalement à la distinction entre la mémoire dite « déclarative » ou « explicite », c’est-à-dire une mémoire qui dépend de l’hippocampe et qui est relative à des souvenirs pouvant être rappelés ou verbalisés consciemment, et la mémoire non-déclarative ou implicite.
L’autre observation inattendue fut que H.M. avait totalement perdu le souvenir des évènements relatifs aux quelques années précédant l’opération mais qu’il se souvenait parfaitement de son enfance. Ce phénomène, appelée amnésie rétrograde partielle, a conduit les chercheurs à émettre l’hypothèse selon laquelle l’hippocampe serait nécessaire à la formation des souvenirs. Selon eux, les souvenirs seraient stockés pendant une période limitée dans l’hippocampe avant d’être transférés* au niveau du cortex pour un stockage à très long terme. Cette phase est appelée la consolidation systémique.
Ces études ont permis de montrer que l’hippocampe, une toute petite partie du cerveau est nécessaire pour l’apprentissage et la mémoire. Il n’est toutefois pas possible de conclure définitivement qu’il s’agit du site même de celle-ci. En effet, cela nécessiterait de connaitre le substrat neuronal de la mémoire, ce qui est encore hors d’atteinte. Il est communément admis que l’apprentissage et la mémoire sont dus à la modification des connexions entre les neurones, plus spécifiquement à l’augmentation du poids des synapses qui permettent aux neurones de communiquer entre eux. Ce processus est appelé plasticité synaptique, mais les règles exactes régissant cette plasticité sont encore mal connues.
Le sommeil et la mémoire
L’hypothèse selon laquelle le sommeil est important dans la mémoire date du début du siècle. Elle a depuis été confirmée par la recherche expérimentale. De nombreuses études ont ainsi montré qu’un apprentissage modifiait les oscillations du cerveau pendant le sommeil. De plus, il a été démontré qu’un apprentissage suivi d’une phase de sommeil était plus efficace, alors qu’une privation de sommeil après un apprentissage diminuait la performance mnésique. Pourtant, il nous est toujours difficile de comprendre pourquoi une tâche spécifique est améliorée par le sommeil quand d’autres ne semblent pas affectées. Par ailleurs, le sommeil n’est pas un bloc uniforme. En effet, on observe d’une part le sommeil paradoxal (appelé sommeil « REM » en anglais) pendant lequel l’activité électrique du cerveau ressemble à celle de l’éveil. Ce type de sommeil est associé aux rêves ayant un contenu riche. Et d’autre part, on observe le sommeil à ondes lentes, qui correspond à des phases de sommeil séparées en trois temps avec un ralentissement progressif de la fréquence des oscillations, associé à une augmentation de leurs amplitudes. Certains résultats suggèrent que le sommeil à ondes lentes serait bénéfique pour la mémoire déclarative dépendant de l’hippocampe, alors que le sommeil paradoxal serait plutôt impliqué dans la mémoire non-déclarative ou des mémoires ayant une forte implication émotionnelle. Cette hypothèse semble très attractive par son apparente simplicité, mais rentre pourtant en contradiction avec certaines observations. Il reste donc beaucoup de questions sans réponse et des conclusions définitives ne pourront survenir que lorsque l’on aura identifié avec précision les mécanismes neuronaux en jeu.
Les deux hypothèses de l’effet bénéfique du sommeil sur la mémoire
Deux grandes théories ont été proposées pour expliquer les effets bénéfiques du sommeil dans la mémoire. Selon la première hypothèse, appelée « homéostasie synaptique (SHY en anglais) », le sommeil serait important pour diminuer le poids des synapses. En effet, comme décrit précédemment, la mémoire est supposée se former dans le cerveau par la modification du poids des synapses qui permettent aux neurones de communiquer entre eux. Ce processus est appelé potentialisation à long terme (PLT). Lors de l’apprentissage, le poids des synapses impliquées dans cet apprentissage est augmenté alors que celui des autres synapses reste inchangé voire diminué. Or, la communication synaptique consomme une grande partie de l’énergie nécessaire au fonctionnement du cerveau ; l’apprentissage et la mémorisation pendant l’éveil s’accompagnent donc d’une forte augmentation de la consommation énergétique. Le sommeil en diminuant le poids des synapses (dépression synaptique), permettrait, par conséquent, de réaliser de grandes économies d’énergie. Cette théorie très élégante permet ainsi de relier à la fois mémoire et apprentissage, ainsi que sommeil et récupération physique.
La deuxième théorie propose que les informations encodées pendant l’éveil soient rejouées pendant le sommeil et que ces réactivations jouent un rôle crucial pour consolider la mémoire et le transfert des souvenirs de l’hippocampe au cortex (consolidation systémique). Cette théorie permet à la fois d’expliquer les effets bénéfiques du sommeil, qui dans certains cas, augmente la capacité de mémorisation, et de mieux comprendre comment les souvenirs sont progressivement transférés de l’hippocampe au cortex.
Alors que pendant très longtemps, l’hypothèse SHY n’accordait aucune importance aux réactivations du sommeil, une version plus récente propose que toutes les synapses voient leur poids diminuer sauf celles qui sont impliquées lors des réactivations. Ces dernières auraient donc pour rôle de protéger certaines synapses de la dépression synaptique, afin de ne pas oublier les souvenirs importants. Selon la deuxième hypothèse, les synapses réactivées seraient au contraire associées à un renforcement du poids synaptique pour consolider les souvenirs, mais également pour les transférer au niveau de cortex pour un stockage à long terme.
La controverse porte donc sur le type de plasticité ayant lieu pendant le sommeil : dépression ou potentialisation synaptique. Il est encore impossible de trancher définitivement entre les deux puisque d’un point de vue théorique, il est extrêmement difficile de démontrer l’absence d’un phénomène. De plus, d’un point de vue technique, il n’est pas encore possible de mesurer directement le poids des synapses d’un grand nombre de neurones. La controverse risque donc de durer encore longtemps, même si de nombreux chercheurs pensent que les deux types de plasticité peuvent parfaitement coexister permettant à la fois de renforcer les souvenirs importants et d’effacer les autres.
Les réactivations du sommeil
Si les deux théories restent opposées sur le type de plasticité à l’œuvre pendant le sommeil, elles reposent toutes les deux sur la nécessité des réactivations du sommeil. Comment ces réactivations ont-elles été observées ? L’influence du sommeil sur la mémoire a trouvé ses lettres de noblesses grâce à l’étude de la mémoire spatiale chez le rongeur. A l’aide des développements technologiques apparus dans les années 90, l’étude de la mémoire spatiale a pu être réalisée à travers l’analyse de l’activité de grands ensembles de neurones chez l’animal libre de ses mouvements.
Les recherches se sont notamment focalisées sur l’activité des neurones hippocampiques (appelés « cellules de lieu »). Ces neurones sont actifs (on dit qu’ils déchargent) lorsqu’un animal occupe un lieu particulier de l’environnement (appelé champ de lieu) (O’Keefe & Nadel, 1978). Cette découverte a valu à son auteur le prix Nobel de médecine en 2014. Comme souvent en science, cela s’avère en réalité plus compliqué. En effet, si chaque neurone ne codait que pour un seul lieu, il faudrait un nombre considérable de neurones pour couvrir tous les environnements que l’on connaît et ce codage serait particulièrement inefficace. Le codage est en fait réalisé par un ensemble de neurones donné, on parle d’assemblée cellulaire. Il s’agit d’un codage combinatoire en population qui est si précis que l’on peut parfaitement prédire la position d’un animal dans un environnement uniquement en regardant l’activité électrique de quelques dizaines de neurones.
Mais quel est le rapport entre ce codage spatial et le sommeil ? C’est en fait grâce à lui que l’on a pu identifier les réactivations du sommeil. Si on place un animal dans un couloir et qu’il réalise toujours la même trajectoire, les cellules de lieu codant pour les différentes parties du couloir seront activées toujours avec la même séquence, selon l’ordre de la position des champs de lieu dans ce corridor. Or, les chercheurs ont observé que les neurones étaient réactivés dans le même ordre durant le sommeil qui suivait l’apprentissage. Ces réactivations étaient observées pendant le sommeil à ondes lentes au moment d’oscillations très rapides et de courte durée appelé « ripples ». Ces observations étaient en accord avec l’hypothèse émise par Gyorgy Buzsaki en 1989, selon laquelle les informations étaient encodées pendant l’éveil et étaient rejouées et consolidées pendant le sommeil, principalement au moment des ripples, ces réactivations agissant comme la répétition dans un apprentissage classique.
Un problème persiste pourtant. En effet, ce n’est pas parce que l’on observe des réactivations neuronales que celles-ci sont vraiment nécessaires. Pour démontrer leur rôle, il faudrait supprimer sélectivement les réactivations du sommeil pour perturber la mémorisation. Or l’identification en temps réel des réactivations est techniquement difficile. Pour contourner ce problème, avec Gabrielle Girardeau et Michael Zugaro au Collège de France, en collaboration avec Gyorgy Buzsaki, nous avons tiré profit du fait que les réactivations ont lieu principalement pendant les ripples. Nous avons ainsi détecté les ripples en temps réel et utilisé une stimulation électrique pour supprimer toute l’activité neuronale de l’hippocampe associée, empêchant par conséquent toutes les réactivations. Les rats devaient résoudre une tâche de mémoire spatiale sur un labyrinthe radial, pour retrouver de la nourriture placée dans seulement 3 des 8 bras du labyrinthe. Les stimulations étaient déclenchées chaque jour, pendant l’heure de sommeil qui suivait l’apprentissage. La suppression des ripples a entrainé une diminution de la performance des rats démontrant ainsi que les ripples participent activement à l’apprentissage.
Création de souvenir artificiels dans la mémoire : une démonstration de l’existence des réactivations
Si cette expérience démontrait le rôle des ripples dans la consolidation de la mémoire, le lien avec les réactivations n’était qu’indirect. Pour s’attaquer à cette question, mon équipe de recherche à l’ESPCI a tenté de faire l’inverse : au lieu de perturber la mémoire, nous avons tenté d’utiliser les réactivations du sommeil pour créer un souvenir artificiel grâce à une interface cerveau-machine. Le but de cette interface cerveau-machine est d’associer la décharge d’une « cellule de lieu » de l’hippocampe avec une stimulation électrique du circuit de la récompense pendant le sommeil. En d’autres termes, nous avons essayé de donner une récompense pendant que l’animal « rêvait [1]» ou plus précisément lorsqu’il « rejouait » un endroit précis. Pour ce faire, nous avons utilisé des stimulations du faisceau médian prosencéphalique (MFB), connue pour induire une forte récompense chez le rongeur et fréquemment utilisé comme renforcement positif dans des tâches comportementales.
Au moyen de l’interface cerveau-machine développée, nous avons associé les réactivations spontanées d’une cellule de lieu pendant le sommeil à une stimulation de récompense, afin de créer artificiellement une préférence de lieu pour l’endroit représenté par celle-ci. Au réveil les souris allaient effectivement dans le champ de lieu du neurone utilisé, démontrant ainsi la possibilité́ de créer des souvenirs artificiels pendant le sommeil. Le temps passé dans le champ de lieu était augmenté significativement alors que le temps mis pour rentrer dans le champ de lieu pour la première fois était diminué. De plus, cet effet n’était visible que dans les premiers essais après le réveil. Comme aucune récompense n’était présentée au réveil de l’animal, le fait qu’il se dirige directement dans le champ de lieu du neurone stimulé suggérait que l’animal s’y rendait pensant y retrouver une récompense. Ceci correspond en fait à une procédure d’extinction classiquement utilisée pour définir un comportement dirigé vers un but (l’animal fait une action pour obtenir une récompense et cesse de faire cette action quand l’action n’est plus récompensée). L’association cellule de lieu-récompense s’est donc manifestée au réveil au niveau comportemental par une association lieu-récompense. Cela démontrait que les cellules de lieu détenaient la même information spatiale durant la navigation et durant le sommeil et donc l’existence des réactivations neuronales pendant le sommeil. Cela mettait également en évidence le fait qu’elles avaient bien un rôle causal dans la navigation spatiale alors que leur rôle n’était jusqu’à présent montré que par des études corrélatives.
Enfin l’utilisation du souvenir artificiellement créé dans un comportement dirigé vers un but nous a permis de prouver qu’il était possible de générer des souvenirs explicites durant le sommeil. Certaines études avaient en effet montré que l’on pouvait créer des conditionnements simples pendant le sommeil, mais dans toutes ces études, l’apprentissage correspondait à l’apparition d’une réaction réflexe à un stimulus. Dans notre cas, aucun stimulus n’était présenté au réveil, l’animal se déplaçait librement mais « choisissait » d’aller dans le champ de lieu. L’ensemble de ces observations montre qu’il est possible de créer des souvenirs artificiels explicites pendant le sommeil. Plus précisément, il est possible de changer la valence émotionnelle d’un souvenir pendant le sommeil : le rendre plus positif ou potentiellement plus négatif.
Quelles applications chez l’homme ?
Une des applications possibles de cette technologie serait le développement de nouvelles thérapies pour soigner le stress post-traumatique. Une des thérapies utilisées actuellement consiste à replacer artificiellement les patients dans la situation traumatique tout en y associant un contexte apaisant pour casser la réaction pathologique de peur. Cependant, quand les patients doivent repenser au souvenir traumatique dans le but de modifier leur valence émotionnelle, ils le font dans des conditions qui ne permettent pas le rappel exact de la trace mnésique de l’événement traumatique. En effet, le patient sait que ce rappel n’est pas réel, mais plus encore, le cerveau lui-même ne se laisse pas tromper, ce qui constitue l’une des hypothèses du relatif manque d’efficacité de ces thérapies. Si l’on pouvait identifier les réactivations des souvenirs traumatiques pendant le sommeil, nous pensons que cela correspondrait au moment optimal pour pouvoir supprimer l’association pathologique avec la réaction de peur générée par l’expérience du traumatisme. Cette modification est plus à même de réussir que pendant l’éveil car le cerveau du patient la considère alors comme bien réelle.
L’ensemble des résultats et des théories présentés ici a abouti au développement de protocoles permettant d’améliorer l’effet bénéfique du sommeil chez l’homme. Il a ainsi été montré que la manipulation de certaines oscillations du sommeil augmentait l’effet bénéfique du sommeil sur la mémoire. La recherche fondamentale cherche maintenant à approfondir les connaissances sur les mécanismes fins mis en jeu pendant le sommeil et la recherche appliquée à améliorer concrètement les effets bénéfiques du sommeil sur la mémorisation et à le rendre accessible au plus grand nombre. Peut-on améliorer les performances des sportifs pendant leur sommeil ? Peut-on limiter les méfaits des maladies neurodégénératives ? Autant de questions découlant de ces travaux auxquels les chercheurs tentent d’apporter des réponses.
[1] Il n’est rigoureusement pas possible de parler de rêve chez l’animal puisqu’on ne peut pas démontrer que l’animal a une expérience consciente pendant son sommeil. Nous pouvons simplement conclure que les cellules de lieu d’un endroit particulier sont actives pendant le sommeil, comme si l’animal rejouait cette expérience. DE plus, les réactivations au moment des ripples sont observées pendant le sommeil à ondes lentes et pas pendant le sommeil paradoxal ou la majeure partie de rêves sont observés.
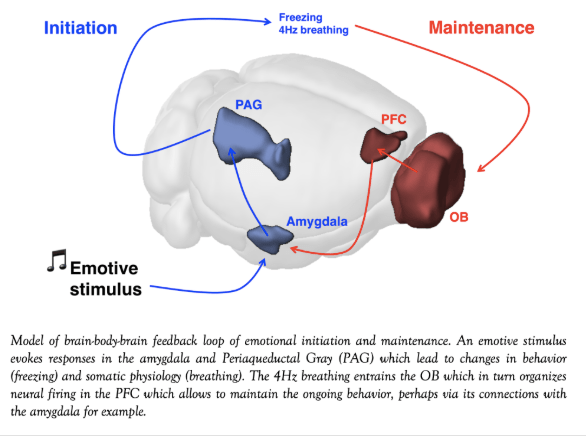

 During my thesis, I studied the role of a protease involved in thrombolysis (destruction of blood clot) the tissue-type plasminogen activator (tPA), which is so far the only available treatment for stroke in human (Benchenane et al. Trends Neurosci 2004). But tPA is also capable of cleaving the NR1 sub-unit of the NMDA receptor leading to an increase in calcium influx into the neurons that enhance ischemic neuronal death (Nicole et al., 2001, Fernandez-Monreal et al., J Biol Chem 2004).
During my thesis, I studied the role of a protease involved in thrombolysis (destruction of blood clot) the tissue-type plasminogen activator (tPA), which is so far the only available treatment for stroke in human (Benchenane et al. Trends Neurosci 2004). But tPA is also capable of cleaving the NR1 sub-unit of the NMDA receptor leading to an increase in calcium influx into the neurons that enhance ischemic neuronal death (Nicole et al., 2001, Fernandez-Monreal et al., J Biol Chem 2004). Memory is fundamental to human life. It allows us to acquire and use information to adapt our behavior in response to experience. Memory is not a unitary concept but is composed of multiple memory types that involve different neuroanatomic structures (Squire, 2004). The memory of facts and events, called declarative memory has been shown to rely on the hippocampus for its formation and temporary storage. But in its initial form, the memory trace is still vulnerable to interference. Thus memory traces, initially labile, are remodeled to become durably stored and retrievable. This process has been called memory consolidation. It occurs in two successive periods. First, the trace is stabilized within the hippocampus; a phase that has been called “cellular consolidation” mainly because several cellular and molecular pathways that mediate this consolidation have been discovered. This has been related to the long-term modification of synaptic weight of particular synapses (increase: long-term potentiation (LTP) or decrease: long-term depression, (LTD)). Interestingly, all the treatments that affect LTP also affect long-term memory making LTP the mostly likely cellular correlates of long-term memory. But this is not the final stage, indeed the memory trace undergoes a second step of consolidation by being transferred in neocortical sites. This has been called “systemic consolidation” since it relies on communication between different brain structures.
Memory is fundamental to human life. It allows us to acquire and use information to adapt our behavior in response to experience. Memory is not a unitary concept but is composed of multiple memory types that involve different neuroanatomic structures (Squire, 2004). The memory of facts and events, called declarative memory has been shown to rely on the hippocampus for its formation and temporary storage. But in its initial form, the memory trace is still vulnerable to interference. Thus memory traces, initially labile, are remodeled to become durably stored and retrievable. This process has been called memory consolidation. It occurs in two successive periods. First, the trace is stabilized within the hippocampus; a phase that has been called “cellular consolidation” mainly because several cellular and molecular pathways that mediate this consolidation have been discovered. This has been related to the long-term modification of synaptic weight of particular synapses (increase: long-term potentiation (LTP) or decrease: long-term depression, (LTD)). Interestingly, all the treatments that affect LTP also affect long-term memory making LTP the mostly likely cellular correlates of long-term memory. But this is not the final stage, indeed the memory trace undergoes a second step of consolidation by being transferred in neocortical sites. This has been called “systemic consolidation” since it relies on communication between different brain structures.




{kind=link}